Chromatin Structure
DNA packaging controls which genes are accessible to RNA polymerase, transcription factors, and regulatory complexes.
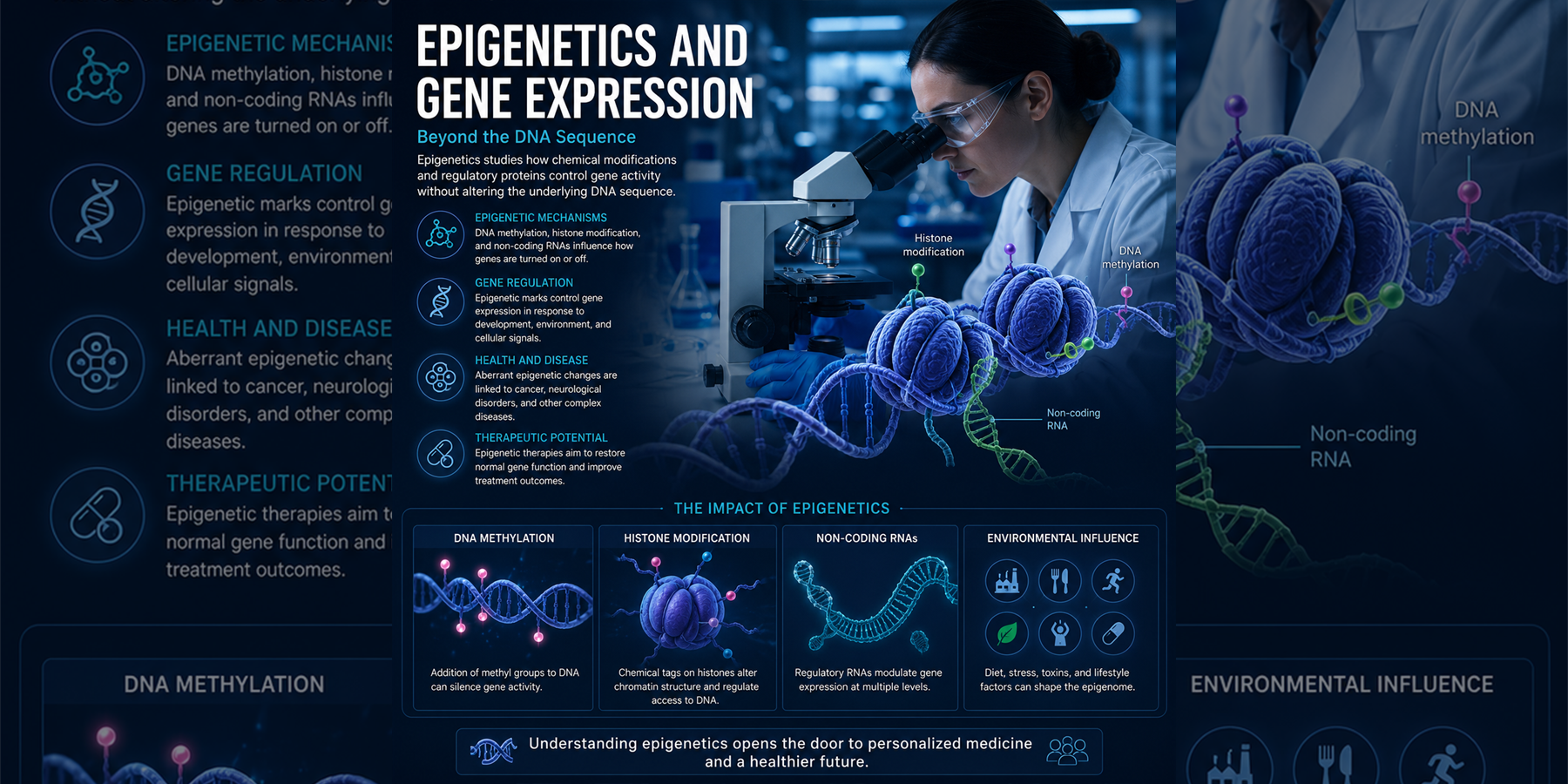
DNA Methylation | Histone Modification | Chromatin | Non-Coding RNA
Molecular mechanisms regulating the genome beyond DNA sequence - governing cellular identity, development, disease, and the response to environmental influences throughout life.
Abstract
Epigenetics studies heritable and reversible changes in gene activity that occur without changing the DNA sequence. These mechanisms shape cellular identity, differentiation, aging, environmental adaptation, and disease.
DNA packaging controls which genes are accessible to RNA polymerase, transcription factors, and regulatory complexes.
CpG methylation can compact chromatin, silence promoters, stabilize cell identity, and suppress repetitive elements.
Acetylation, methylation, and remodeling marks create a dynamic regulatory code read by effector proteins.
miRNAs, lncRNAs, and siRNAs regulate transcripts, chromatin states, genome defense, and nuclear architecture.
Cancer, neurological disorders, autoimmune disease, metabolic conditions, and aging all involve altered epigenetic control.
DNMT, HDAC, EZH2, and IDH inhibitors show how epigenetic mechanisms can become therapeutic targets.
Part I
Different cell types share nearly identical genomes yet produce radically different structures and functions. Epigenetic regulation resolves this biological paradox.
Part II
The molecular machinery reads the genetic code and converts it into functional proteins that govern cellular activity.
Core DNA sequences upstream of the transcription start site where RNA polymerase II assembles.
Distal regulatory elements boost transcription by looping to contact gene promoters across large genomic distances.
DNA-binding proteins recruit co-activators and basal transcription machinery to regulate gene output.
These elements repress, restrict, or boundary enhancer activity to preserve correct cell-type expression.
Part III
The 3D packaging of DNA into chromatin is a major regulatory layer that controls gene accessibility.
About 147 base pairs of DNA wrap around a histone octamer made of H2A, H2B, H3, and H4 proteins.
Flexible N-terminal tails protrude from nucleosomes and carry post-translational modifications.
Loosely packed chromatin is transcriptionally permissive, enriched in H3K27ac and H3K4me3 marks.
Dense chromatin maintains silencing of repetitive elements, centromeres, and inappropriate developmental genes.
Part IV
The addition of methyl groups to cytosine residues is one of the most extensively studied epigenetic mechanisms.
CpG island methylation recruits methyl-CpG binding proteins and repressive complexes, compacting chromatin and blocking transcription.
Dynamic methylation reprogramming orchestrates cell fate decisions during embryogenesis and tissue specification.
Methylation of repetitive elements silences transposons, SINEs, and LINEs to protect genome integrity.
Lineage-specific methylation patterns are maintained through cell division as epigenetic memory.
Insulin-like growth factor that promotes fetal growth.
Long non-coding RNA involved in tumor suppression and growth regulation.
XIST lncRNA coats the inactive X chromosome and recruits silencing machinery.
Part V
Post-translational modifications on histone tails form a dynamic regulatory code that controls gene activity.
Histone acetylation neutralizes lysine charge, loosens chromatin, and supports transcriptional activation.
Marks active enhancers and promoters and is used to map enhancer activity.
Associated with active gene bodies and transcriptional elongation.
Decondenses chromatin fiber and counters heterochromatin spreading.
Associated with active promoters and transcription-ready genes.
Associated with heterochromatin, repetitive element silencing, and compact chromatin.
Polycomb-mediated repression of developmental genes until activation is required.
ATP-dependent complexes reposition, eject, or restructure nucleosomes to alter DNA accessibility.
Effector proteins read histone marks while enzyme complexes add or remove regulatory modifications.
Part VI
Most of the human genome is transcribed into RNA, much of it non-coding but functionally critical for gene control.
miRNAs bind target mRNAs and recruit RISC to drive mRNA degradation or translation repression.
Examples: miR-21, let-7 family, miR-155.
lncRNAs guide chromatin modifiers, scaffold protein complexes, and organize nuclear compartments.
Examples: XIST, HOTAIR, NEAT1.
siRNAs trigger mRNA degradation and can support transcriptional silencing through chromatin changes.
Examples: transposon silencing, antiviral defense, Inclisiran.
Part VII
A single fertilized egg generates more than 200 specialized cell types through orchestrated epigenetic programming.
Global reprogramming erases parental marks; paternal demethylation precedes passive maternal dilution.
Inner cell mass retains pluripotency through Polycomb repression and Oct4, Sox2, Nanog networks.
Three germ layers establish distinct epigenetic landscapes and poised bivalent chromatin domains.
Cell-type enhancers activate through transcription factor binding, H3K27 acetylation, and chromatin looping.
H3K4me3 and H3K27me3 coexist at developmental promoters, enabling rapid activation or repression.
Lineage-specific enhancers acquire H3K4me1 before full activation.
DNMT1, Polycomb complexes, HP1, CTCF, and cohesin help restore cell identity after replication.
Part VIII
Epigenetic dysregulation is a fundamental driver of cancer, neurological, autoimmune, and metabolic diseases.
Cancer cells simultaneously activate oncogenes and silence tumor suppressors through aberrant methylation and histone modification.
Global hypomethylation, promoter hypermethylation, H3K27me3 reprogramming, and IDH mutations that create CpG island methylator phenotypes.
Epigenetic mechanisms are critical in the brain, where gene regulation supports learning, memory, and neuronal circuit function.
Alzheimer's disease, Parkinson's disease, autism spectrum disorders, and Rett syndrome.
Aberrant regulation of immune cell identity contributes to dysregulated lymphocyte activation and loss of immune tolerance.
Lupus, rheumatoid arthritis, and multiple sclerosis.
Environmental and nutritional inputs modulate epigenomes in metabolically active tissues.
Type 2 diabetes, obesity, and cardiovascular disease.
Part IX
Environmental exposures reshape epigenomes throughout life, and epigenetic therapies are transforming oncology.
Folate, methionine, and B vitamins are methyl group donors affecting global methylation.
Glucocorticoid signaling can modify HPA axis methylation and stress response programming.
Heavy metals, air pollution, and endocrine disruptors alter methylation and histone marks.
Exercise induces transient demethylation at muscle enhancers and alters circulating miRNA profiles.
Epigenetic clocks estimate biological age through CpG methylation changes and genome-wide drift.
DNA-demethylating agents used in MDS and AML to reactivate silenced tumor suppressors.
Promote histone acetylation and can induce differentiation or apoptosis in cancer cells.
Target methyltransferase or oncometabolite pathways to restore normal differentiation programs.
Part X
Emerging technologies are reshaping epigenomics research, cancer diagnostics, regenerative medicine, and precision healthcare.
scATAC-seq, bisulfite sequencing, and CUT&TAG reveal cell-to-cell epigenetic heterogeneity.
Spatial assays map chromatin accessibility and methylation within intact tissue context.
dCas9 fused to DNMT3A, TET1, p300, or KRAB rewrites gene activity without cutting DNA.
Deep learning predicts chromatin state, transcription factor binding, and gene expression from sequence.
Partial reprogramming with Yamanaka factors may reverse epigenetic aging marks in tissues.
Methylation-based liquid biopsy can support multi-cancer early detection and tissue-of-origin prediction.
Scientific References
Alberts, B., Johnson, A., Lewis, J., et al. (2022). Molecular Biology of the Cell (7th ed.). Garland Science.
Allis, C. D., Caparros, M. L., Jenuwein, T., Reinberg, D., & Lachner, M. (2015). Epigenetics (2nd ed.). Cold Spring Harbor Laboratory Press.
Bird, A. (2007). Perceptions of Epigenetics. Nature, 447(7143), 396-398.
Goldberg, A. D., Allis, C. D., & Bernstein, E. (2007). Epigenetics: A Landscape Takes Shape. Cell, 128(4), 635-638.
Jaenisch, R., & Bird, A. (2003). Epigenetic Regulation of Gene Expression: How the Genome Integrates Intrinsic and Environmental Signals. Nature Genetics, 33, 245-254.
Lodish, H., Berk, A., Kaiser, C. A., et al. (2021). Molecular Cell Biology (9th ed.). W.H. Freeman.
Moore, L. D., Le, T., & Fan, G. (2013). DNA Methylation and Its Basic Function. Neuropsychopharmacology, 38(1), 23-38.
Ptashne, M. (2013). Epigenetics: Core Misconcept. Proceedings of the National Academy of Sciences, 110(18), 7101-7103.
Riggs, A. D., Martienssen, R. A., & Russo, V. E. A. (1996). Epigenetic Mechanisms of Gene Regulation. Cold Spring Harbor Laboratory Press.
Roadmap Epigenomics Consortium. (2015). Integrative Analysis of 111 Reference Human Epigenomes. Nature, 518(7539), 317-330.
Takahashi, K., & Yamanaka, S. (2006). Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell, 126(4), 663-676.
FAQ
Evidence-based answers to the most common questions on epigenetics and gene expression.
Epigenetics is the study of heritable changes in gene expression that do not alter the underlying DNA sequence. Major mechanisms include DNA methylation, histone modification, chromatin remodeling, and non-coding RNA regulation.
DNA methylation adds methyl groups to cytosine bases, often at CpG sites, to regulate promoter activity, repetitive element silencing, imprinting, X-chromosome inactivation, and cellular identity.
Histone marks change chromatin accessibility and recruit reader proteins that activate or repress gene expression depending on the modification and genomic context.
Many epigenetic marks are copied during cell division, allowing daughter cells to inherit transcriptional programs. Some marks can also be influenced by developmental or environmental exposure.
Cancer often involves global hypomethylation, promoter hypermethylation of tumor suppressors, histone modification changes, and mutations in epigenetic enzymes.